
Graphidaceae of Central French Guiana
Rebecca Yahr
Glossary
of technical terms
By H. Sipman, Botanic Garden & Botanical Museum Berlin-Dahlem
Rebecca Yahr
Glossary
of technical terms
By H. Sipman, Botanic Garden & Botanical Museum Berlin-Dahlem
This survey of the Graphidaceae is a first step towards producing a lichen florula of central French Guiana and will be completed in several stages, starting with keys, photos and descriptions of the species with transversely septate spores in the genera Phaeographis, Sarcographa and Graphis. The next stage will comprise the rest of the Graphidaceae, including Graphina and Phaeographina. The material described here was collected by Richard Harris and Bill Buck during three trips between 1990 and 1997, when over 2,000 collections of lichenized fungi were made near the village of Saül in central French Guiana. I am also grateful for the chance to examine the collections of Philip May, gathered in 1997 from central French Guiana. His material contains several taxa not included here.
The Graphidaceae are commonly understood to consist of eight genera. Because there has been no phylogenetic approach to delimiting genera in this group, these genera are conveniently if artificially defined on the basis of spore morphology and development of stromatic tissue (differentiated, loosely woven tissue within which ascomata are immersed). A genus has been erected for each combination of hyaline or brown, transversely-septate or muriform spored, non-stromatic groups (Graphis, Phaeographis, Graphina, Phaeographina) and stromatic groups (Glyphis, Sarcographa, Meduselina, Sarcographina). Although these boundaries are artificial and individual taxa are well-known to incorporate grades of each, it is a convenient system of classification until the systematics of the family is better understood. It is worth noting here that this work is intended as a descriptive floristic project and cannot address the substantial systematic work that remains in the family.
Previous reports (before 1987) from the Guianas have been summarized by Hekking and Sipman (1988), who, after accounting for synonymies, include three varieties of Glyphis, 17 species of Graphis, four species of Phaeographis, and seven species of Sarcographa from French Guiana. For comparison from Brazil, Redinger (1934, 1935) reports two species of Glyphis (although G. triseptata Redgr. should likely be considered another variant of G. confluens), 48 species of Graphis (G. illinata Escwh. has been transferred to Graphina illinata (Eschw.) Hale), 17 species of Phaeographis and five species of Sarcographa. Taking only the genus Graphis, to attempt to understand the extent of overlap in collections or the differences in taxonomy, I have compared which taxa are known from each region according to different workers. Only seven of Redingers reported taxa were also found in French Guiana as of 1987. This treatment brings that total number up to eight. However, of the material examined, of the 17 taxa known from French Guiana before 1987, I have found only three.
This inconsistency may be indicative of several factors: 1) geographic limitations, since the material examined for this treatment was all collected in and around one region in the vicinity of Saül, 2) lack of habitat diversity sampled, since the majority of specimens were collected in lowland moist forest, on notably shady and humid tree trunks, 3) taxonomic or systematic inconsistencies between workers concepts of taxa. I am unable to address the geography or the taxonomic issues. However, I can cursorily address the lack of diversity of sampled habitats. Most of the collections were made on tree trunks in the low elevation, humid and shady forest. These understory habitats are well differentiated ecologically from the exposed branches of the canopy or from higher elevation exposed forest, where sunlight is more frequent and intense and humidity fluctuates more widely and regularly. The lichen floras that correspond to each habitat can similarly be expected to differ widely. Representative collections of canopy habitats was accomplished through collections from fallen twigs where possible; in addition, one collecting trip was made to a more exposed forest type on higher elevation rock outcrops. These habitats were by no means sampled at the same intensity as those of moist forests. Nonetheless, it is immediately apparent that the flora of the canopy is quite separate from that in the understory. It is likely that with further examination of the canopy collections, more taxa will be added to this treatment. In a cursory glance through the fallen canopy collections, about half were not yet seen by me. Although a few collections from the canopy are included here, the coverage is far better for understory species.
Keys and Key Characters
The keys are artificial and make use of characters that are almost certainly
unfamiliar and sometimes somewhat ambiguous to most non-specialists. However,
with a brief introduction to the ways these descriptive terms and character
states are used, it is easy to gain a working understanding of the group sufficient
to use keys here and in other relevant literature. I will attempt to describe
their expected ranges of variation and comment on their taxonomic utility below.
 The
first of these to understand is spore morphology. This is a character whose
value is somewhat suspect on the generic level although it is consistently --
although not entirely satisfactorily used to define generic limits. There
are well known cases in the family of so-called "sporomorphs",
where one taxon can be clearly circumscribed based on a whole suite of morphological
and chemical characters, but can be found with either transversely-septate or
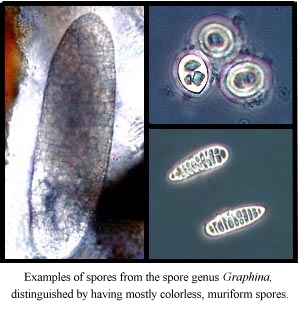
muriform spores (spores with both transverse and longitudinal septae). In these
keys, I will follow the artificial breakdown of spore septation to lead the
user to the same taxon using both transversely septate or muriform keys. Therefore
a few Graphina species will be found in the
Graphis key and vice versa. In addition, there are several instances
in which a transversely-septate spored species is occasionally or even commonly
found with one or two longitudinal septae, but
is retained in the genus Graphis. These are also found in both the transversely-septate
and muriform-spored keys.
The
first of these to understand is spore morphology. This is a character whose
value is somewhat suspect on the generic level although it is consistently --
although not entirely satisfactorily used to define generic limits. There
are well known cases in the family of so-called "sporomorphs",
where one taxon can be clearly circumscribed based on a whole suite of morphological
and chemical characters, but can be found with either transversely-septate or
muriform spores (spores with both transverse and longitudinal septae). In these
keys, I will follow the artificial breakdown of spore septation to lead the
user to the same taxon using both transversely septate or muriform keys. Therefore
a few Graphina species will be found in the
Graphis key and vice versa. In addition, there are several instances
in which a transversely-septate spored species is occasionally or even commonly
found with one or two longitudinal septae, but
is retained in the genus Graphis. These are also found in both the transversely-septate
and muriform-spored keys.
Likewise, "brown" and "colorless" do not adequately represent the range of variation present in spore coloration. Many senescent or deteriorating spores attain some level of pigmentation, but these can often be distinguished since they are wrinkled and generally sickly-looking. In this treatment (and in most), truly brown spores in the transversely-septate taxa are easy to distinguish since they are also fairly consistently thick-walled, symmetrical and oblong. These will belong in either Phaeographis or Sarcographa depending upon stromatic development. In the muriform spored taxa, a whole grade of hyaline through yellowish to brownish can be found. I have tried to make the keys as straightforward as possible, dividing hyaline from not hyaline and describing the taxa in whichever genus they are currently recognized.
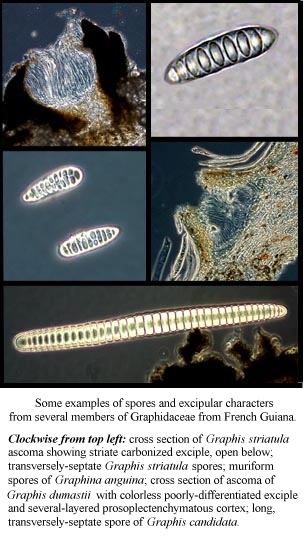
The exciple in the Graphidaceae is an important character at the species level and historically on the subgeneric level. It is the modified layer of tissue that directly surrounds the hymenium and can be quite variable in the graphids overall. Two important distinctions are made in the keys regarding this layer of tissue: degree of carbonization and presence or absence of striation. Carbonization ranges from a thick, continuous band of black, brittle tissue completely enclosing the hymenium to dusty inclusions of moderately-darkened tissue at the tips of the labia. The most important distinction in the keys is whether carbonization is present, and if so, whether or not it is continuous or closed below the hymenium. In rare cases, the only carbonized tissue is below the hymenium. The degree of carbonization can be difficult to observe and is best assessed with thin cross sections of ascomata, all the way down to the bark. If unsure, include any dark brown areas as carbonized on your first run through the key but be careful to make sections as thin as possible since a thick slice of pale tissue can appear darkened or black under the microscope!
Striae may or may not be continuous along the length of most ascomata and are variable in thickness, abundance and developmental stage. It is obvious that narrowly-defined states of this character are of questionable use in rigid definitions of taxa; however, some taxa tend to have striae and some do not. Within those that do, they are often best observed on older (usually larger overall) lirellae and can often be seen in cross section under the dissecting scope at high magnification or in a water mount of a thin section. Generally, the thinner the section, the better this character can be observed. It is best to make sections of several lirellae before deciding whether or not striae are present. Striae can consist of carbonized or not-carbonized excipular tissue or can be made of crystals. Wirth and Hale (1978) made careful observations of this character and discuss it in detail. They mounted all specimens in Hoyers and lightly boiled them to evacuate any air spaces which can obscure color differences. However, I have found that this is not necessary to observe the excipular characters used in this treatment (or in theirs).
Wirth and Hale (1978) contend that undue emphasis has been placed on both carbonization and striation, since each can be variable within the same thallus. This position is also justifiable based on my observations. Nonetheless, until a monographic treatment can be completed using many specimens of each type, it will be impossible to determine just how much consistency in these characters can be expected.
Another apparently useful character is the presence or absence and consistency of the covering over the ascomata, if it is distinct from the thallus. This tissue is referred to here as the thalline veil and can be comprised of anything from a powdery or mealy thin covering of hyphae, to a thick, continuous, cartilaginous corticate layer, with or without algae and/or crystals. Some are brightly pigmented (as in the brilliant carmine veil of Graphis miniata Redgr.); if the exciple below the veil is carbonized, it is often possible to see the black areas through the veil where it has been rubbed away or worn thin. Fissurine taxa without well-differentiated exciples are not considered to have a distinct covering since the ascomata are imbedded in the thallus.
Thallus characters are not very reliable, as colors can be variable depending on age and condition of thallus, whether or not it has been grazed, etc. One character that is reliable however, is the presence of a layered, clear prosoplectenchymatous cortex. This is easily seen in cross section as a colorless layer with coherent hyphae and which peels away in thin sheets. Since it is sometimes quite friable, it may be lost in thin sections unless care is taken to preserve it. Presence or absence of a cortex overall is usually a good character as well. Where grazing or damage has removed it in places, it is usually possible to tell that this is the case. Ecorticate specimens look very finely powdery or mealy and occasionally encrusted with oxalate crystals (as in Graphis koreaiensis Sipman).
Chemically, most of the Graphidaceae contain either no lichen substances or one or two of a limited set of other compounds: norstictic acid (sometimes with satellites of hypoconstictic, constictic, or connorstictic), stictic acid (sometimes with satellites of hypoconstictic, constictic, or connorstictic), psoromic acid, protocetraric acid or lichexanthone. Many contain abundant inclusions of calcium oxalate crystals in the thallus, often concentrated near the ascomata and sometimes within the exciple. In addition, many spores of the Graphidaceae are reactive with I and with KI. With only I, many spores will turn a dark blue-violet color or a lighter smoky purple-gray. In some species, with KI, the margins of the hymenium stain light blue. However, some taxa have no reactions with I nor with KI. The chemistry of the taxa presented here was tested using TLC with a benzene-dioxane-acetic acid solvent system and silica gel, aluminum backed precoated plates.
The keys often contain unnamed specimens which have been referred to by their collection number. All specimens are deposited at NY.
Special thanks go to Scott Mori for his interest and enthusiasm in supporting this and other botanical studies on central French Guiana; to Dick Harris for immeasurable assistance in the form of experience and advice on the Graphidaceae thoughtfully given and for unsurpassed hand-sections for photography; to Bill Buck for cheerfully offered inspiration and organizational and administrative assistance; and to David Goldstein, Anita Kahn and family for friendship and housing during my stay in New York.
References
Redinger, K. 1934. Die Graphidineen der ersten Regnellschen Expedition nach Brasilien, 1892-94, II: Graphina und Phaeographina. Arkiv för Botanik. Band 26A No.1: 1-105.
Redinger, K. 1935. Die Graphidineen der ersten Regnellschen Expedition nach Brasilien, 1892-94, III: Graphis und Phaeographis, nebst einem Nachtrage zu Graphina. Arkiv för Botanik. Band 27A. No. 3: 1-103.
Wirth, M. and M. E. Hale, Jr. 1978. Morden-Smithsonian Expedition to Dominica: The Lichens (Graphidaceae). Smithsonian Contributions to botany, No. 40: 1-64.